
Karl Edlinger
Abenddämmerung des Darwinismus – brauchen wir eine neue Evolutionstheorie? *
Fassen wir das Wesentliche der gewonnenen Erkenntnisse zusammen,,so betrifft dies zunächst eine Korrektur der gewohnten Naturauffassung. Es ist nicht mehr das Bild eines Kampfes alles Lebenden untereinander, sondern das Bild eines stillen, ausgeglichenen Tributfriedens. Jeder Organismus zahlt seinen Tribut an jene spezialisierten Tiere, die ihm von der Natur als Regulatoren zugewiesen sind, die ihn von dem Überschuß an Nachkommenschaft befreien müssen, der ansonsten seinen Lebensraum überfüllen würde. Das Tributzahlen aber ist eine Quantitäts- und keine Qualitätsfrage. Den Tribut zahlen jene Individuen, die ihr Unstern in den Weg eines Feindes fährt. Dieser nimmt, was ihm begegnet, ohne Feinauswahl nach Strich oder Farbton. Damit stimmt gut, daß die Tierwelt von heute keineswegs aus lauter Bestangepaßtem besteht, sondern eine unerschöpfliche Mannigfaltigkeit bunter, phantastischer Gestalten umfaßt, die augenfällig das Gepräge zweckloser Schönheit tragen. Die Tiere haben keine „Schutzmittel“, die Regulatoren werden nicht „abgewehrt“, sondern finden reichlich Nahrung, die ihrem Spezialgeschmack voll entspricht. Daß die Tiere nicht aussterben, ist darin begründet, daß sie so viel Nachkommenschaft produzieren, daß der Ausfall durch organische und unorganische feindliche Einflüsse gedeckt ist und außerdem genug Individuen zur Fortpflanzung kommen, um die Art in annähernd gleicher Stärke weiterzuführen.
Franz Heikertinger 1954
Einleitung
Seit bald eineinhalb Jahrhunderten, seit dem Erscheinen der Erstauflage von Charles Darwins „Origin of Species….“ scheint die darwinistische Evolutionstheorie in einem unaufhaltsamen Siegeszug begriffen, der sie zu einem der am wenigsten bestrittenen und kritisierten Gedankengebäude unserer Zeit machte. Widerstand gegen das, was allgemein unter dem Kürzel „Darwinismus“ begriffen wird, scheint sich nur mehr in religiös fundamentalistischen Kreisen wie etwa im „Bibelgürtel“ des Amerikanischen Südens zu regen und zu formieren, ansonsten genießt der Darwinismus in seiner heutigen Form, nämlich als Neodarwinismus oder Synthetische Theorie der Evolution, eine Akzeptanz und Immunität gegen jede Kritik, die an religiöse Dogmen vergangener Epochen erinnert.
Charles Darwins Werk (1809-1882) figuriert in einem allgemeinen, vereinfachten Bild der Wissenschaftsgeschichte gleichrangig mit dem eines N. Kopernikus oder I. Newton als einer jener Meilensteine, die nur in größeren Abständen die geistigen Revolutionen markieren, welche die Menschheit jeweils um ein gewaltiges Stück vorwärts brachten, andererseits aber auch eine Revision des Bildes bedeuteten, das sich die Menschen von sich selbst und ihrer Stellung in der Natur machten und den Menschen so auch von so manchem selbsterrichteten Sockel stürzten.
Kritischen Geistern geben gerade solche Situationen, solch unangefochten dominierende Weltbilder oder Theoriengebäude, zu denken. Es fragt sich, ob das Bild von Charles Darwin und seinem Werk tatsächlich der Realität entspricht und vor allem, inwieweit die Geltungsansprüche und das Interpretationsmonopol des Darwinismus vergangener Zeiten sowie seiner modernen Form tatsächlich berechtigt sind. Auf Grund sehr eindruckvoller historischer Vorbilder sollte die Berufung auf Autoritäten in dieser Frage nicht den Ausschlag geben.
Historisches
Beleuchtet man die geschichtliche Entwicklung näher, so drängt sich sofort eine Unterscheidung auf, ohne die der Werdegang des Evolutionsdenkens insgesamt und des Darwinismus in all seinen Spielarten nicht verständlich sind, nämlich zwischen der Überzeugung vom allmählichen Wandel der Lebewesen an sich und der Konsequenz, dass sich die Organismen aus anorganischen Materialien allmählich herausbildeten einerseits und verschiedenen Erklärungsmustern dafür auf der anderen Seite.
Der Wandel der Organismen wurde lange vor Darwin von verschiedenen Naturwissenschaftlern mit unterschiedlicher Radikalität vertreten, etwa von Charles Bonnet, Georges Buffon, Jean Baptiste Lamarck, Erasmus Darwin (dem Großvater von Charles Darwin), Leopold von Buch, Edward Blyth, Geoffroy de Saint Hilaire oder Herbert Spencer, um nur einige zu nennen. Charles Darwin empfing von ihnen zahlreiche Anregungen, die aber in der Literatur weitgehend ignoriert werden. Vor allem der fundamentale Beitrag Edward Blyths wird konsequent übersehen oder geleugnet (Mayr 1984).
Nur durch weitgehendes Ignorieren der Vorläufer konnten jene Mythen entstehen, die sich immer noch um Darwin ranken, etwa der von der wütenden Ablehnung seines Hauptwerks oder vom genialen Einfall während des Aufenthalts auf den Galapagosinseln und der Beobachtung der dortigen Finken. Die Realität wirkt hier ernüchternd, denn erstens war der Boden für das Evolutionsdenken längst bereitet, vor allem durch J. B. Lamarcks Werk, und zweitens kommt man bei einiger historischer Sorgfalt nicht an dem jüngeren Zeitgenossen Darwins, Alfred Russel Wallace, vorbei, der aber ähnlich wie beider Vorläufer lange ignoriert wurde, obwohl er zeitgleich mit Darwin, manche vermuten, vor ihm (Glaubrecht 2002) , entscheidende theoretische Schritte in Richtung der heute dominierenden Erklärung der Evolution machte.
Sowohl Darwin als auch Wallace gehen in Anlehnung an die Schriften von T. R. Malthus davon aus, dass normalerweise in jeder Art von Generation zu Generation mehr Nachkommen hervorgebracht werden, als zur Ersetzung der sterbenden Individuen nötig wären. Unter diesen Nachkommen, welche untereinander variieren und sich auch gegenüber den Eltern teilweise unterscheiden, findet ein ständiger Konkurrenzkampf um Energie, Nahrung, Sexualpartner, Raum u. v. a. m. statt, in dem die bestgeeigneten Individuen überleben. Sie erweisen sich solcherart als besser an ihre Lebensbedingungen oder Umwelt angepasst und haben größere Chancen, sich fortzupflanzen und ihre Eigenschaften und Merkmale an die Nachkommen weiterzugeben. Für das Überleben günstige Eigenschaften verstärken sich so, ungünstige werden allmählich eliminiert. Arten verändern sich so durch ein Wechselspiel von Variation und Selektion, unter Anpassung an die Anforderungen der Umwelt.
Diese Sicht wird allgemein, etwas verkürzt, als die Darwins vorgestellt, sie ist es aber nur bedingt. Im Gegensatz zu Wallace, der seine eigene Theorie gleichzeitig mit Darwin vorstellte, postuliert Darwin in Anlehnung an Lamarck und ähnlich wie Herbert Spencer zusätzlich zum Mechanismus von Variation und Selektion auch eine aktive Anpassung der Organismen an die Umwelt und die Möglichkeit einer Vererbung von solcherart erworbenen Merkmalen. In seinem Spätwerk über das Variieren der Pflanzen und Tiere (Darwin 1868) versucht er diese Sicht sogar durch eine eigene „Keimchenhypothese“ zu begründen, die allerdings 1875 von seinem Vetter Francis Galton experimentell widerlegt wurde.
Der Neodarwinismus (Ultradarwinismus)
Wallace könnte also mit Recht als der erste echte Darwinist gelten. Er teilt sich diesen Titel aber mit August Weismann, der mit seiner Keimbahnhypothese (Weismann 1893), in der einer eigenen, noch nicht näher umrissenen, aber von gerichteten Umwelteinflüssen weitgehend unabhängigen Substanz die ausschließliche Rolle bei der Vererbung zugesprochen wurde.
Weismann wurde damit zum Begründer des modernen Neodarwinismus oder Ultradarwinismus, der unter Verzicht auf Darwins lamarckistische Postulate die durch Variation und Selektion begründete Anpassung hypostasierte. In dieses Gedankengebäude wurden schließlich die klassische und die molekulare Genetik sowie fast sämtliche übrigen Teilgebiete bis zur vergleichenden Anatomie und Morphologie integriert. In Anlehnung an die Mutationslehre von de Vries, dem man aber in seiner Sicht von Mutationen als große umwälzende erbliche Veränderungen nicht folgte, stellte sich Evolution als durch das Wechselspiel von Mutation und Selektion verursachtes Anpassungsgeschehen der lebendigen Natur dar.
Verschiedene Beobachtungen führten zu Erweiterungen des adaptationistischen Konzepts, vor allem zeigen populationsdynamische Prozesse und die Untersuchung von Inselfloren oder Inselfaunen, dass bei dauernder Isolation weniger Individuen einer Art oder bei Wanderungen die Veränderungs- und Artbildungsprozesse beschleunigt werden können. Dieses Konzept geht auf den heute unbekannten Moriz Wagner zurück, der sich kritisch von Darwin distanzierte und zu Lebzeiten vor allem von August Weismann heftig angegriffen wurde.
Nun zeigt aber die kritische Untersuchung der Literatur, dass die Beispiele für darwinistische Anpassung immer nur Einzelmerkmale betreffen, deren Wandel in den meisten Fällen reversibel ist, also gar nicht als Beleg für unumkehrbare Evolutionsprozesse gelten kann. Geradezu als Paradebeispiele für diese „darwinistische“ Anpassung wurden und werden immer wieder die schon von Wallace oft angeführten zahlreichen Fälle von Tarnung Mimese und Mimikry angeführt.
Tarnungs- und Mimikryhypothesen wurden unter vielen anderen durch Moriz Wagner (1868-1889) oder durch den heute wohl zu Unrecht vergessenen Wiener Entomologen Franz Heikertinger (1954) kritischen Analysen unterzogen, die in dem durch eingehende Diskussionen aller Für und Wider begründeten Schluss gipfelten, dass sie jeder ökologischen Evidenz und damit auch des Anpassungswerts im darwinistischen Sinne entbehren. Tarn- und Warntrachten der Tiere funktionieren als solche ebenso wenig wie Schutzsubstanzen und andere angeblich durch stammesgeschichtliche Anpassung entstandene Einrichtungen.
Die Bedrohung, die von einer solchen Sicht ausgehen könnte, scheint unterschwellig mitzuwirken, wenn in der Literatur über stammesgeschichtliche Fragen ein inflationärer Gebrauch des Begriffs Anpassung um sich greift (vgl. v. Mayr 1991). Im Herder Lexikon der Biologie (Sauermost 1994, 1. Bd. s. 191) werden Anpassungen als Eigenschaften bezeichnet, die den Organismen die „lebenserhaltende Auseinandersetzung mit den verschiedensten Umweltbedingungen gestatten“. Angepasstheit wird oft als Lebensfähigkeit schlechthin definiert, was die Sinnhaftigkeit des Begriffs in Frage stellt.
Diese Zweifel kommen in einer Arbeit zum Ausdruck, die der weltberühmte Paläontologe Steven J. Gould zusammen mit Richard C. Lewontine verfasste und in der die beiden den Nachweis führten, dass die Anpassungsmetapher in ihrer Logik mit dem Argument des Dr. Pangloss in Voltaires Candide vergleichbar sei. Dr. Pangloss war davon überzeugt, in der besten aller möglichen Welten zu leben und fand sogar angesichts der nahenden persönlichen Katastrophe noch den Mut, diese als notwendig für einen guten Verlauf der Weltgeschehens zu bezeichnen. Ganz ähnlich argumentiert der Doyen der Synthetischen Theorie Ernst Mayr (1991), wenn er fordert, dass man immer dann, wenn eine adaptationistische Begründung für ein Merkmal widerlegt sei, eben nach einer anderen Anpassungsbegründung suchen müsse.
Der Adaptationismus mündet so in einen Dogmatismus, der die Frage nach seiner generellen Berechtigung aufwirft. Immerhin haben seine Kritiker das Argument für sich, dass Organismen, wenn man die oben erwähnte vage Definition für Angepasstheit zugrundelegt, prinzipiell nur angepasste Organismen existieren können, da wer nicht lebensfähig ist, eben nicht leben kann. Wie solche angebliche Anpassungen aber zustande kommen, kann mit diesem theoretischen Rüstzeug nicht geklärt werden. Vor allem gelingt es nicht, große Linien des organismischen Wandels adaptationistisch zu begründen (Reichholf 1993).
So verwundert es nicht, wenn das Dogma der darwinistischen Anpassung als Motor der Evolution von manchen Biologen wie Gould & Lewontin /1979), Reichholf (1993, 1997, 2004) oder Peters & Peters (1997) vorsichtig, von anderen wie Heikertinger (1954) oder Edlinger, Gutmann & Weingarten (1991) radikal in Frage gestellt wird.
Physikalistische Metaphern statt Organismen
Zudem zeigte sich in den letzten Jahrzehnten ein weiteres Problem: Das Fehlen eines konsistenten, aus der Biologie stammenden und Lebewesen einigermaßen adäquat definierenden Organismusbegriffs- und Modells.
Die Konsequenz ist, dass in der Literatur in zunehmendem Ausmaß Organismusmetaphern Platz greifen, die entweder aus anderen naturwissenschaftlichen Disziplinen, vor allem aus der Physik, stammen, oder überhaupt das Resultat von Spekulationen bzw. Computerspielen darstellen. In solchen Modellen stellen sich dann Lebewesen, soweit auf ihre Struktur überhaupt eingegangen wird, bestenfalls als inkohärente Molekülwolken, passiv durchflossen von Materie und Energie, dar.
Ähnliche Probleme wirft der molekularbiologische Reduktionismus auf, der Organismen letztlich auf Genarrangements reduziert.
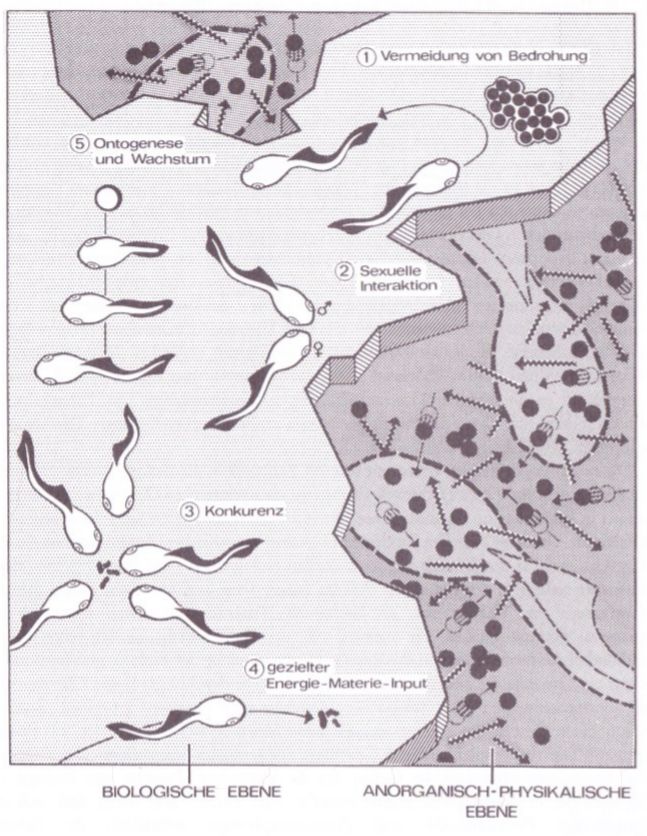 Abb. 1) Physikalistische Organismusmodelle (rechts, dunkles Feld) im Gegensatz zu denen der unten behandelten Organismischen Konstruktionslehre (links, helles Feld), welche von den grundlegenden Eigenschaften der Lebeweesen ausgehen.
Abb. 1) Physikalistische Organismusmodelle (rechts, dunkles Feld) im Gegensatz zu denen der unten behandelten Organismischen Konstruktionslehre (links, helles Feld), welche von den grundlegenden Eigenschaften der Lebeweesen ausgehen.
Die Organismische Konstruktionslehre
Vor mehr fast Jahrzehnten traf sich in Frankfurt am Main eine Runde von jüngeren Biologen des Senckenberg-Instituts, die sich ein sehr anspruchsvolles Ziel gesetzt hatten. In Frankfurt waren damals die Studentenunruhen der späten sechziger Jahre zu Ende gegangen und die Umbruchsstimmung, die sie erzeugt hatten, war noch in aller Erinnerung. Eine „Kritische Theorie“ sollte nach dem Willen der revolutionären Studenten und ihrer akademischen Mentoren, wie etwas Theodor Adorno oder Herbert Marcuse ein neues Bewusstsein von sozialen Entwicklungen und ihren ideologischen Implikationen schaffen und die verknöcherten und rückschrittlichen Strukturen der Gesellschaft insgesamt aufzubrechen.
Von diesem Elan beeindruckt begannen die Frankfurter Biologen, die verknöcherte Strukturen auch in der Zoologie und Botanik, vor allem aber in der Evolutionslehre bemerkten, die Frage der Evolutionsmechanismen (die Evolution als solche wird nicht in Frage gestellt) neu zu stellen und über Lösungen nachzudenken, die vor allem aus den bereits erwähnten Sackgassen und Erkenntnisfallen der darwinistischen Biologie herausführen sollten. Die Situation war ihrem Vorhaben günstig. Am Senckenberg-Institut, benannt nach seinem Begründer, dem Humanisten, Philanhthropen und Goethe-Freund Johann Christian Senckenberg, weht traditionell ein liberaler Wind und der damalige Direktor Wilhelm Schäfer war selber auf der Suche nach neuen Methoden der Darstellung lebender Organismen, welche die bislang übliche Fragmentierung, die schon von J. Schaxel (1922) kritisierte Auflösung der Lebewesen in ihre Teile vermeiden sollten.
Die Grundlage für eine ganz eigene, ganzheitliche, organismische Theorie der Lebewesen war damit gelegt. Anfangs noch etwas holprig formuliert und in vieler Hinsicht noch mit Restbeständen der alten bekämpften Theorien behaftet, entwickelte sich die „Frankfurter Lehre“, in deutlichem Anklang an die andere im aufgeschlossenen geistigen Klima Frankfurts geborene Erneuerung auch „Kritische Evolutionstheorie“ genannt, allmählich zu dem, was heute als Organismische Konstruktionslehre bekannt ist.
In zahlreichen Publikationen wurde der theoretische Fortschritt dokumentiert, der schließlich dazu führte, auf die Anpassungsvorstellung zu verzichten und ein Organismuskonzept vorzulegen, das nicht nur die Autonomie der Lebewesen begründet, sondern auch aufzeigt, wie die anderen naturwissenschaftlichen Disziplinen ohne theoretische Ungereimtheiten in biologische Theorien intergriert werden können (Gutmann & Bonik 1981; Gutmann & Edlinger 1994a, b; Edlinger & Gutmann 2002, Gutmann & Edlinger .2002, Gudo 2004).
Dabei wird von Prinzipien ausgegangen, die für Lebewesen spezifisch sind und durch die gesamte Organismenwelt als Grundinvarianten hindurchgereicht werden und Gültigkeit beanspruchen.
Technische Metaphern
Die Grundgedanken sind relativ einfach und anhand von Analogie mit der Technik gut verständlich zu machen. Man stelle sich Autos vor, die technischen Geräte, die uns allen am geläufigsten sind. Autos unterscheiden sich voneinander in vielen äußerlichen Merkmalen, sie sind unterschiedlich groß, schnell und stark, haben einen unterschiedlichen Energieverbrauch und sind auch für sehr verschiedene Verwendungen gebaut. Äußerliche Merkmale sind den Gestaltmerkmalen von Pflanzen und Tieren oder auch ihren Farben vergleichbar. Größe, erreichbare Geschwindigkeit und kW-Leistung entsprechen der Konstitution von Lebwesen, der Energieverbrauch möge der Energieverwertung der Organismen entsprechen und der Zweck den ökologischen Beziehungen, also den Umweltkontakten, welche die Lebewesen eingehen können.
Wer nun glaubt, die Eigenschaften eines Autos allein aus den Anforderungen erklären zu können, die ihre beabsichtigte Verwendung, also ihre Umwelt an sie stellt, der macht sich die Sache wohl zu einfach und geht vor allem von völlig falschen Voraussetzungen aus.
Beginnen wir beim äußeren Erscheinungsbild! Dieses ist in vieler Hinsicht von Geschmacksfragen abhängig, die beim Entwurf und der technischen Planung von Autos eine große Rolle spielen. Käufer orientieren sich ja sehr häufig an solchen Nebensächlichkeiten und nicht ausschließlich an technischen Details. Sicher kann man, wie die Autos bestimmter Hobby-Techniker zeigen, das Äußere eines Pkw zwar ins Groteske umkehren, doch nur unter einer Bedingung: Die Funktionstüchtigkeit muss gewährleistet bleiben. Und damit sind dem freien Gestaltungswillen Fesseln angelegt.
Vor allem betrifft dies den Aufbau und die Anordnung des Herzstücks jedes Kraftfahrzeugs, des Motors. Es gibt zwar zahlreiche verschiedene Arten von Motoren, doch ist ihnen allen gemeinsam, dass sie Energie von einer chemischen Form in mechanischen Antrieb umwandeln. Dies erfordert eine reibungslose Zufuhr des Triebstoffs, eine problemlose Vergasung, Verdichtung und Zündung in geschlossenen Zylindern, in denen sich wiederum Kolben möglichst reibungsarm bewegen, um ihrerseits wieder eine Kurbelwelle anzutreiben, die den mechanischen Antrieb weiterleitet. Die Kurbelwelle selber ist wiederum nur dann sinnvoll, wenn die Zylinder in einer ganz fein abgestimmten Ordnung „zusammenarbeiten“.
Ebensolche Feinabstimmung ist vom Getriebe gefordert, und dass die Räder an bestimmten Stellen dort und so montiert sein müssen, dass über Getriebe, Kardanwelle und verschiedene andere Übertragungsmechanismen die Kraft an sie weitergegeben werden, scheint selbstverständlich, stellt aber wiederum eine unumgehbare Einschränkung und Festlegung dar.
Ebenso ist die Anordnung des Auspuffs, des Kühlsystems, der Lichtmaschine und anderer Teile nicht beliebig, sondern hängt eng mit ihrer Funktion zusammen.
Wir können also feststellen, dass es bei Autos, ebenso wie bei anderen Maschinen, bestimmte Konstruktionszwänge gibt, die dem freien Gestaltungswillen des Technikers sehr enge Grenzen setzen.
So wie wir in unserem Auto-Beispiel denkt nun die Organismische Konstruktionslehre in vieler, beileibe nicht in jeder Hinsicht, über die Lebewesen.
Auch sie können ebenso wie Autos als kraftschlüssige Maschinen betrachtet werden, in denen Energie gewandelt, von chemisch gebundener Form in mechanische, in Bewegung umgesetzt wird. So wie die einzelnen Bestandteile eines Autos sind die Bestandteile des Organismus, seine Zellen, Gewebe, Muskeln, Sehnen, Darm und andere Organe oder bei Pflanzen die verschiedenen Leitbündel und anderen Teile in einer präzise abgestimmten Ordnung arrangiert und können dadurch weitgehend problemlos funktionieren.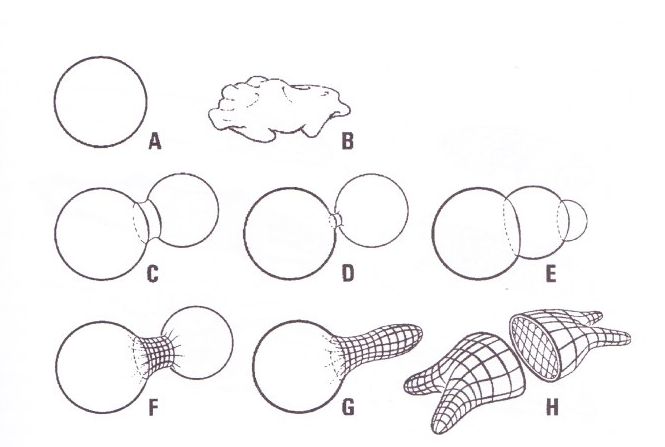
Abb. 2) Im Rahmen der Konstruktionstheorie stellt sich Form als Ergebnis von Prozessen, als Ausdruck weiterlaufender Prozesse mit vorabgelaufener permanenter Sicherung der Form oder als erstarrtes Resultat von Prozessen (Skelette) dar. Jede Formgebung ist nur über einen Genese-Prozess und die Bedingungen der präzedenten Organisation zu begründen. Diese Aussage gilt auf allen Ebenen der Organisation, trifft für alle übergreifenden oder in sich kraftschlüssigen Subeinheiten zu.
Operationale Geschlossenheit der Membran, Kohärenz, d.h. mechanische Verbindung und Verkoppelung aller wirksamen Teilstrukturen, ist für die Erklärung der Formerzeugung und der Formveränderung zwingende Voraussetzung. Ein entkoppeltes oder nur teilweise festgelegtes Element wird zum Teil der Füllung und kann nicht an der Formerzeugung oder Bewegungserzwingung teilnehmen; Öffnung der Membran führt zum Auslaufen und zum Kollaps der Verspannungen; Abkoppelung von Teilen der Membran oder Unvollständigkeit der Vernetzung oder Lückenhaftigkeit in der Abspannung der Membran führt in den nicht restringierten Bereichen zur kugeligen Aufwölbung.
A: Normalform eines intern nicht verspannten Hydrauliksystems. Diese Form ist das Ergebnis der Interaktion von osmotischer Expansionstendenz und Spannung in der geschlossenen Membran.
B: jede kollabierte Form wird zum Spielball äußerer Kräfte. Ob eine Form Ergebnis äußerer Kräfte ist (bei Unterfüllung) oder Resultat dichter Verspannung, kann nicht durch visuelle Inspektion erkannt werden, sondern muss durch morphologische Prüfung ermittelt werden.
C-D: Einschnürung zwischen zwei sich teilenden Zellen.
F, G: Teilweise Bandagierung sichert nur begrenzte Formkontrolle, die unverspannten Bereiche nehmen Kugelform an.
H: Total formkontrollierte Zellgestalt, die durch Abspannung in allen Hauptrichtungen des Raumes gesichert ist. Membran und formzügelnde Fibrillen müssen ein mechanisch kohärentes System bilden.
Die Spontaneität und Autonomie, d. h. die grundsätzliche Eigenständigkeit gegenüber der Außenwelt und der spezielle Antrieb ihrer Tätigkeit von innen, der zum Unterschied von Autos niemals erlahmt, werden später und aufbauend auf der Konstruktion zu erklären sein. Doch erst einmal zu den grundsätzlichen Aussagen der Organismischen Konstruktionslehre!
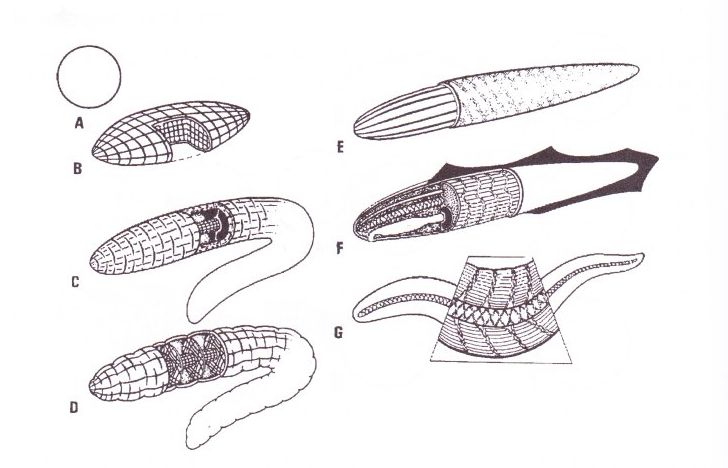
Abb. 3) A: Automatisch erreichte Form bei nicht aktiver oder fehlender interner Abspannung.
B: Abflachung (wie bei Plattwürmern, Nemertinen etc.) kann nur durch enge dorsoventrale Elemente geschehen. Abspannung in allen Raumrichtungen legt die Form insgesamt fest.
C: Wurmgestalt mit einheitlichem flüssigkeitsgefülltem Innenraum. Der Körperschlauch mit Längs- und Ringmuskeln muss runde Querschnittform annehmen, sobald die formativen Elemente gespannt sind. Eine Abwandlung des Querschnittes ist nicht möglich.
D: Wurmorganisation mit dicht liegenden Verspannungsstrukturen, in diesem Falle wird Abwandlung des Querschnittes, Abflachung oder laterale Kompression möglich. Die Sicherung der Querschnittform überlagert sich der schlauchförmigen Grundorganisation.
E: Cuticuläre Abspannung in Schlauchform gestattet Auslegung allein mit Längsmuskeln; der Cuticula-Schlauch unterbindet Verkürzung, sobald die Cuticula Fasern bei Anspannung der Muskeln gestrafft sind; von diesem Moment an können nur noch Biegungen durchgeführt werden.
F, G: Bei Sicherung der Längenkonstanz durch eine Chorda-Achse (auch diese besteht aus einem gefüllten Faserstrumpf, der sich durch Verhinderung der Querschnittvergrößerung der Verkürzung widersetzt), ist eine Einhängung der Achse in die Konstruktion nötig. – Myosepten bei Chordaten und Vertebraten.
Die Grundmechanismen der Formbildung sind (bei spezifischer und andersartiger Staffelung der Muskel-, Bindegewebs- und Epithel-Strukturen) die gleichen wie bei einzelnen Zellen. Kohärenz und operationale Geschlossenheit bilden auch auf der Metazoenebene absolute Voraussetzung der Formbestimmung.
Die Kohärenz von Metazoen wird bei Abschluss durch Epithelien vermittels bindegewebiger Vernetzung aller Strukturen erreicht; die Muskeln sind in das fibrilläre Bindegewebsnetz einbezogen, das sie, wie Spannungsglieder wirkend, verstellen, wobei sie über die Füllung wirken. Einzelne Zugrichtungsverläufe können muskulär nicht gesichert sein, sondern sie müssen durch reines Bindegewebe, Basalmembranen oder Cuticulen abgespannt sein.
Die Ausbildung der Form jedes Organismus und aller kohärenten Subsysteme erfolgt hydraulisch, durch expansive osmotisch bedingte Wirkung der Füllung gegen die Spannung der Membran und die Zügelung der Form durch interne Verspannungen oder äußere Bandagen. Jede Form kann in den Mechanismen ihrer Erzwingung nur dann verstanden werden, wenn eine Verpackung der jeweiligen Substanz in sphärischer Form als Bezugspunkt genommen wird. Auch jede ablaufende Bewegung setzt vor ihrem Einsetzen eine aktive Sicherung der Form voraus und erfordert beim Ablauf der Propulsion oder der Erzeugung von jedem anderen Verhalten eine Erhaltung der Form durch Absicherung der Nichtkugeligkeit und durch Unterbindung aller nicht für den Ablauf geforderten Bewegungen.
Jeder Organismus ist nach dieser Theorie konstruktiv zu verstehen. Er ist ein mechanisches, energiewandelndes, hydraulisches System, das durch Wechselwirkung von hydraulischer Füllung und äußerer, meist membranöser Begrenzung primär zur Kugelform tendiert und sich durch selektiv durchlässige Membranen nicht nur gegen die Außenwelt abgrenzt, sondern gerade durch diese selektive Durchlässigkeit mit ihr jene Kontakte herstellen kann, die aus inneren Notwendigkeiten des Organismus heraus eingegangen werden müssen. Niemals geht diese Kontaktaufnahme primär von der Umwelt aus.
Durch innere fasrige Verspannung oder äußere Vergurtung werden von der Kugelform abweichende Formen hergestellt und gesichert. Die Verspannungen und Vergurtungen stehen in einem antagonistischen Verhältnis zur Hydraulik.
Energiewandel
Leben ist energiegetriebenes Geschehen. Organismen arbeiten aus sich heraus. Sie verformen sich durch Kontraktion von Fasern unter Energieverbrauch.
Die zu dieser aus dem Inneren heraus angetriebenen Arbeit nötige Energie wird durch Absorption von (Sonnen-)Strahlung oder Einverleibung von organischer Nahrung und darauffolgende chemische Wandlungsprozesse beschafft. Die Energieversorgung wird aber ebenso aktiv durch den Organismus initiiert, dieser kann also nicht, wie in verschiedenen physikalisch inspirierten Organismustheorien als passiv durch Materie und Energie durchströmt angesehen werden.
Die chemischen Reaktionen, die den Energiewandel ermöglichen und zum Aufbau körpereigener Substanzen nötig sind, sind an die mechanische Konstruktion gebunden. Das heißt, jene Stoffe, die sie überhaupt erst bewirken, die sogenannten Enzyme, sind oft und vor allem wenn sie zusammenwirken sollen, an den Membranen der Zellen und ihrer Bestandteile so angeordnet, dass viele Reaktionen hintereinander ablaufen können. Wenn zum Beispiel Zuckermoleküle abgebaut werden sollen, wird durch ein Enzym einmal eine Spaltung in zwei kleinere Bestandteile bewirkt. Diese werden dann gleichsam an das nächste Enzym „weitergereicht“, das sie dann weiter abbaut. Und so laufen sehr komplizierte chemische Prozesse gleichsam in Reaktions-„ketten“ ab, bis der Abbau abgeschlossen ist. Das selbe gilt für Aufbaureaktionen, also die Synthese verschiedener „körpereigener“ Stoffe aus den Abbauprodukten der Nahrung oder, bei Pflanzen, aus jenen organischen Stoffen, die durch Photosynthese gewonnen werden.
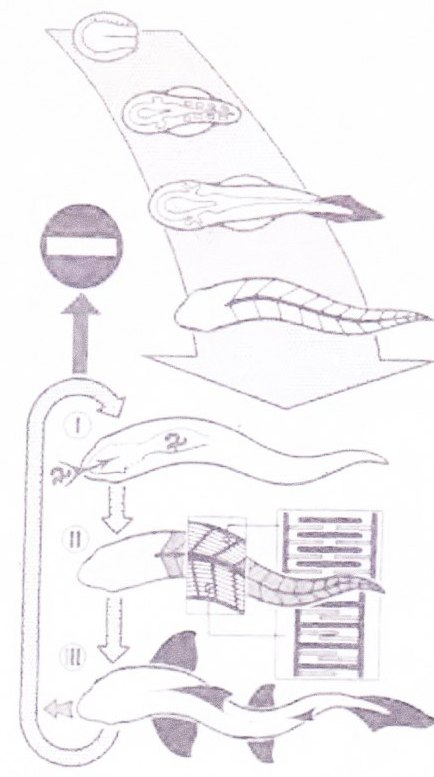 Abb. 4) Alle Lebensaktion besteht aus energetischem Trieb mechanisch kohärenter Konstruktionen; dies gilt für Adultstadien wie Ontogenese in gleicher Weise. Der obere Teil der Abbildung stellt die Embryonal-Entwicklung der Fischkonstruktion dar, die von einer befruchteten Eizelle ihren Ausgang nimmt. Das ontogenetische Geschehen verläuft als energetisch getriebene Aktion eines mechanischen Gebildes, das sich durch Ausweitung von Höhlen und Ausbildung restringierender Strukturen selbst durch interne Aktion ausformt.
Abb. 4) Alle Lebensaktion besteht aus energetischem Trieb mechanisch kohärenter Konstruktionen; dies gilt für Adultstadien wie Ontogenese in gleicher Weise. Der obere Teil der Abbildung stellt die Embryonal-Entwicklung der Fischkonstruktion dar, die von einer befruchteten Eizelle ihren Ausgang nimmt. Das ontogenetische Geschehen verläuft als energetisch getriebene Aktion eines mechanischen Gebildes, das sich durch Ausweitung von Höhlen und Ausbildung restringierender Strukturen selbst durch interne Aktion ausformt.
Jede Formbildung erfolgt hydraulisch durch osmotische Expansion bei membranöser Abgeschlossenheit und formbestimmender Zügelung durch fibrilläre Strukturen im kohärenten Verband. Formbildung stellt Bewegung am Ort dar und als Ausdruck der die Form bestimmenden Kräfte.
Die fertige Konstruktion, ein schematisierter Fisch, arbeitet energiewandelnd, indem aufgenommene Nahrung in Verdauung und Stoffwechsel aufgeschlossen, im Stoffwechsel körperspezifisch umgewandelt (I) als Energiezufuhr für die mechanische Konstruktion genutzt, chemische Energie in mechanische umgesetzt (Ü) und der Körper durch innere Arbeit Propulsion bewirkend deformiert wird, sodass im Rahmen des Bionomiekreises neuer Materie- und Energie-Input (Nahrung) besorgt und in neue Lebensaktion, aber auch in Reproduktion umgesetzt werden kann. Jede Bewegung kann nur auf hydraulischer Grundlage im konstruktiven Verband erfolgen.
Die ebenfalls erforderliche permanente Entsorgung, die natürlich durch mechanische Aktion der Konstruktion bewirkt wird, ist nicht dargestellt, bildet aber mit Selbstversorgung und Reproduktion essentielle Mechanismen im Rahmen der Bionomie.
Die Stellung der „Gene“
Die Enzyme selber und die Botenstoffe, die in Organismen wirksam werden können, haben einen Aufbau, der durch die Erbsubstanz bestimmt ist. Sie enthält gleichsam in verschlüsselter Form die Struktur von Eiweißen, die bei Bedarf erzeugt werden. Das heißt, dass jede Veränderung des chemischen Milieus in den Zellen und Geweben auf die Erbsubstanz wirkt. Diese ist nämlich in komplexen Körpern, den Chromosomen arrangiert. Die Chromosomen haben wieder eine ganz spezielle Struktur, die es erlaubt, auf die „chemischen Signale“, die aus ihrer Umgebung kommen, zu „reagieren“ und jene Abschnitte der Erbsubstanz zu aktivieren, die gerade „gebraucht“ werden, wie es das Operon-Modell von Jacob und Monod zeigt. Steigt zum Beispiel der Zuckergehalt an, dann werden „Gene“ aktiviert, welche die Bildung von zuckerabbauenden Enzymen bewirken. Wenn der Zuckergehalt dann wieder einen „normalen“ Pegel erreicht hat, werden sie wieder „abgeschaltet“. Gene haben also keine hierarchische Sonderstellung, sondern werden ja nach den Notwendigkeiten des Stoffwechsels aktiviert oder abgeschaltet. In vielen Fällen wirken auch mechanische Vorgänge, etwa die Kontraktion von Fasern, dadurch auf die Erbsubstanz, dass sie über Verengung von Zellporen die Zufuhr bestimmter Stoffe verhindern oder aber die Viskosität des Protoplasmas verändern. Auf diese Weise können auch Bewegungen von ganzen Muskeln mittelbar auf die Biochemie der Zellen und Gewebe einwirken. Biomechanik und Biochemie sind also sowohl durch diese Möglichkeit der Einwirkung als auch dadurch untrennbar miteinander verwoben, dass mechanische Bewegung erst durch jene Energie möglich wird, welche durch die biochemischen Abläufe frei wird. Bereiter-Hahn erarbeitete auf der Basis zahlreicher Forschungsergebnisse ein Modell, das der weit verbreiteten Sicht einer hierarchisch übergeordneten Stellung des genetischen Materials (der „genetischen Information“) widerspricht, andererseits aber in die Organismische Konstruktionslehre problemlos integriert werden kann.
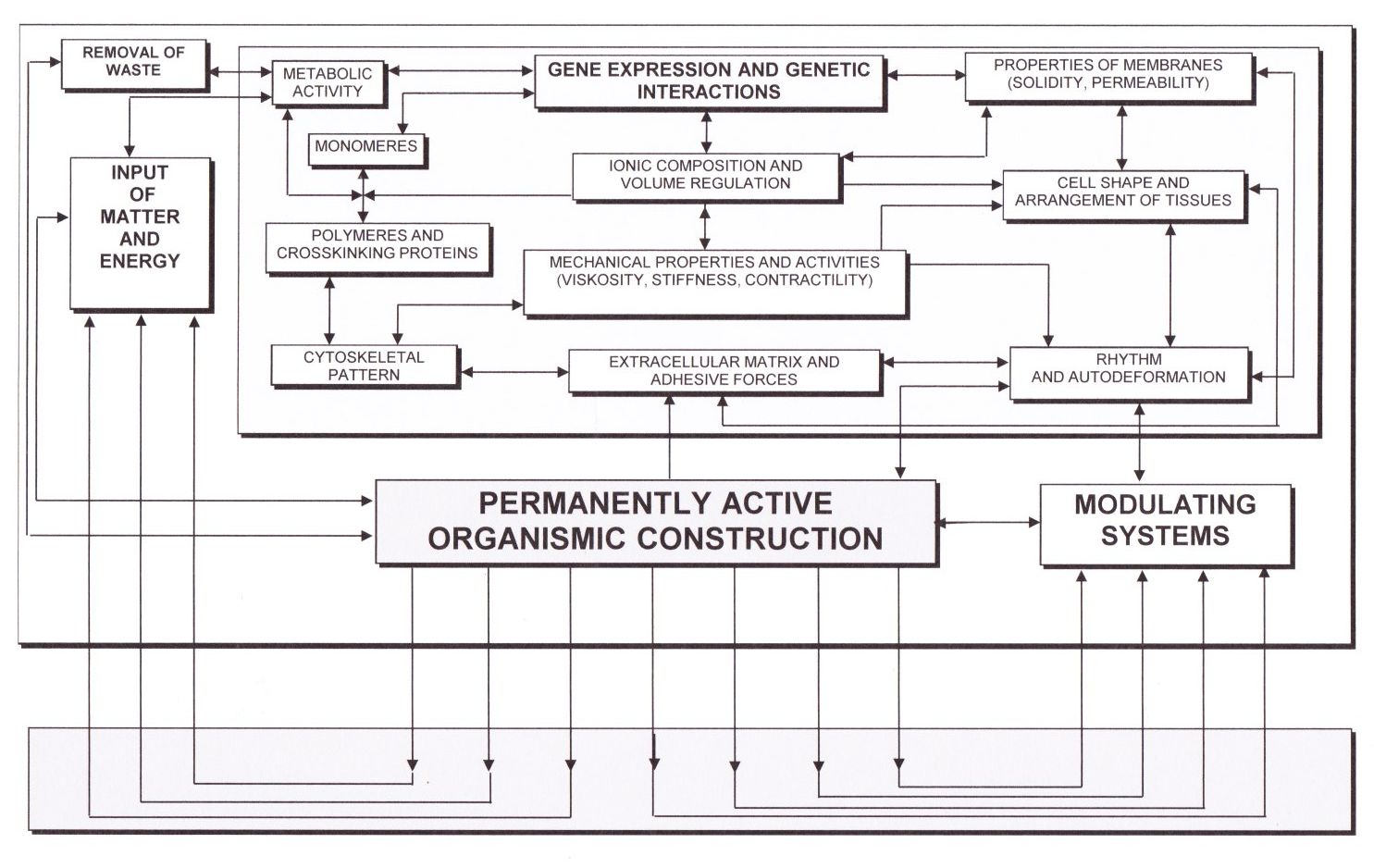
Abb. 5) Ein vereinfachtes Schema der Interaktionsbeziehungen zwischen verschiedenen Komponenten und Funktionen der Zellen, der Organismen und der Umwelt weist den genetischen Mechanismen eine zwar prominente Rolle zu, zeigt aber gleichzeitig, dass sie nur einen Aspekt der mannigfachen innerorganismischen Wechselwirkungen abdecken (Nach Bereiter-Hahn 1991 u. Gutmann & Edlinger 2002).
Rhythmizität
Ständige mechanische Verformung bewirkt innere Zirkulation und Umwälzung von flüssigen, gel- oder soloartigen Bestandteilen und Kraftübertragung auf die Umwelt, also Energieinjektion.
Die Aktivität der Organismen ist immer rhythmisch. Die Rhythmizität ist Resultat wechselseitigen aktivierenden und hemmenden Einflusses von kontraktilen Fasern aufeinander, bei komplexeren Tieren von Nervenzellen.
Nur durch rhythmische, aufeinander abgestimmte Verformung der einzelnen mechanischen Elemente kann sichergestellt werden, dass es zu keinen destruktiv wirkenden Friktionen im Energiewandel, wie etwa Zerrungen und Rissen kommt, dass also das funktionierende Gesamtgefüge unbeschädigt erhalten bleibt.
Alle von den regelmäßigen rhythmischen Verformungen abweichenden Bewegungen stellen Modulationen dar. Sie werden durch teilweise Hemmung der Rhythmik bewirkt.
Reproduktion
Die Organismen sind ständig zur Energieaufnahme bereit, ja ihre innere selbstgenerierte Aktivität besteht im wesentlichen aus Aufnahme und Wandel von Energie. Dadurch entsteht zwangsweise das Bedürfnis, Überangebote, übermäßig aufgenommene Energie abzuleiten und zu entsorgen.
Dies geschieht durch verstärktes Anarbeiten gegen Widerständigkeiten der Außenwelt, durch Wachstum und auch durch reproduktive Proliferation, das heißt Fortpflanzung.
Es gibt viele verschiedenen Möglichkeiten der Fortpflanzung. Sie ist dann möglich, wenn durch Teilung in gleiche oder ungleiche Teile Organismussplitter entstehen, die alle für eine weitere Existenz unabdingbaren mechanischen und chemischen Elemente in einem wenigstens minimal geordneten und friktionslosen Arrangement enthalten.
Hier sind auch Abwandlungen möglich, die sich in der Stammesgeschichte der Lebewesen niederschlagen.
Im Laufe der Stammesgeschichte bildeten sich auch ganz spezielle Fortpflanzungsweisen heraus, bei denen Erbsubstanz in ganz präzise festgelegten Portionen in sogenannten „Keimzellen“ gelagert wird. Solche Keimzellen können dann miteinander verschmelzen und dadurch auch die in ihnen enthaltene Erbsubstanz neu kombinieren. Wir sprechen dann von sexueller Fortpflanzung.
Solche Fortpflanzungsaktivitäten, seien nun sexuelle Stadien zwischengeschaltet oder nicht, haben neues Wachstum zur Folge. Wachstum bedeutet ständige Erzeugung konstruktiver Elemente, die in den Organismus eingebaut werden. Der Einbau erfolgt opportunistisch, d. h. die Mechanik oder den Chemismus hemmende Elemente können jederzeit neu arrangiert oder überhaupt wieder eliminiert werden.
Wachstum bewirkt immer Änderungen im physiologischen Zustand, sei es eine Veränderung der Oberflächen-Volums-Relationen, eine verminderte Arbeitsfähigkeit kontraktiler Fasern oder anderes.
Die dadurch auftretenden Störungen und Friktionen werden durch opportunistische kleine Wandlungsschritte ständig behoben. Diese Wandlungsschritte führen zu Kompensationen und sind notwendigerweise von Veränderungen im Gesamtgefüge begleitet, die aber die Grundinvarianten der hydraulischen, mechanischen Konstruktion nicht verletzen dürfen.
Beispielsweise können Einstülpungsprozesse als Kompensationen für sich verändernde Oberflächen-Volums-Relationen betrachtet werden.
Somit wird auch Ontogenese begreifbar als ständiger durch Wachstumsprozesse vorangetriebener kompensatorischer Wandel, der es dem Organismus ermöglicht, in jeder Phase seiner Entwicklung voll funktions- und lebensfähig zu bleiben.
Kompensation erfordert auch der ständige selbst initiierte Kontakt mit den Widerständigkeiten der Umwelt und das aktive Einschalten in deren Energieströme.
Da ja, soll Bewegung nicht ins Leere laufen, die mechanische Aktivität der Organismen auf sie zurückwirkt, kann die Art des Zurückwirkens durch kompensatorische Veränderung der mechanisch aktiven Strukturen beantwortet werden. Es kommt zu spezifischen Ausformungen des Bewegungsapparats.
Hartskelette
Als energiesparende Einrichtungen können zusätzlich zum Hydroskelett aus verhärtenden Ausscheidungen starre Strukturen entstehen, die den Bewegungsapparat unterstützen bzw. die Kraftübertragung ökonomischer gestalten oder auch als Ansatzflächen für kontraktile Fasern oder Muskulatur dienen. Wir sprechen dann von Innen- oder Außenskeletten (wie etwa Molluskenschalen oder Insektenpanzer).
Nervensysteme
Bei komplexeren Tieren mit zelliger Muskelstruktur etablieren sich schnell besonders differenzierte Muskelzellen, die Nervenzellen, welche mit Muskeln und auch untereinander in Kontakt treten können. Sie sind auf die Generierung elektrischer Erregungsmuster spezialisiert, mit denen sie Muskeln zur Aktivität reizen, aber auch einander erregen oder hemmen können. Sie haben nach wie vor Bewegungs- und Wachstumsfähigkeit und können vor allem in Lückensysteme der organismischen Konstruktion einwachsen. Dabei werden ständig neue (synaptische) Verbindungen zu Muskelzellen geknüpft.
Diese arbeiten ständig antagonistisch gegeneinander und formieren sich so zu geordneten, leistungsfähigen Arrangements.
Durch die erregende Tätigkeit der Nervenzellen wird dieser Formierungsprozess beschleunigt. Dabei werden aber nur solche Nerven-Muskel-Kontakte auf Dauer aufrechterhalten und stabilisiert, welche die rhythmische Aktivität oder die modulierte Bewegungsweise nicht stören. Ansonsten werden sie abgebaut.
Damit etabliert sich durch ständigen opportunistischen Umbau ein sehr effizientes, je nach Komplexität des Tieres vielfältiger modulierbares Erregungs- und Bewegungsmuster.
Durch die ständige Walkung hydraulischer Konstruktionen ist die Bildung größerer Neuronenkonzentrationen, die auch untereinander komplexere Verknüpfungsmuster bilden könnten, begrenzt.
Nur wo große walkungsfreie Hohlräume existieren oder entstehen, können größere Nervenkonzentrationen auftreten. Sie können die unmittelbaren Erregungsmuster des Bewegungsapparats überlagern, stabilisieren aber ihrerseits nur solche Verschaltungsmuster, welche die Mechanik nicht stören und destruieren.
Es zeigt sich so, dass auch die komplexesten Nervensysteme noch von der mechanischen Konstruktion der betreffenden Organismen bestimmt sind.
Neuronen können aber auch Außenstrukturen wie Membranen, Hautareale oder Bewegungscilien ansteuern und zu rhythmischer Aktivität veranlassen. Wird die Rhythmizität durch Widerständigkeiten der Außenwelt oder durch Energieflüsse gestört, kann die modulierende Kompensation durch den Bewegungsapparat des Gesamtorganismus durchgeführt werden. Durch (natürlich nicht bewusstes) Abtesten der verschiedenen Kompensationstätigkeiten auf ihren möglichen Erfolg gewinnen die selbstgenerierten Umweltreize allmählich für den Organismus einen spezifischen Charakter.
Affektion von Cilien durch elektromagnetische Wellen etwa wird allmählich dazu führen, dass nicht der unmittelbare mechanische Einfluss auf ein nahes Objekt gesucht wird, sondern dass sich (primitive) Muster von Entfernungen ausbilden, die auch ein adäquates Agieren ermöglichen. Mechanische Affektion wird die unmittelbare Nähe eines Gegenstandes, den Kontakt mit ihm anzeigen, was sich dann ebenfalls in ganz spezifischen Handlungsabläufen niederschlägt, die allmählich auch fixiert werden (Edlinger 1992a, 2002).
Dazu kommt, dass die Möglichkeit, sich Reizen auszusetzen, durch die organismische Konstruktion und seine Außenstrukturen wesentlich bestimmt und auch beschränkt ist. Dass aber auch, wenn bestimmte Reizmodalitäten oder -qualitäten keine ökonomisch und friktionslos verarbeitbaren Erregungsmuster ermöglichen, auf sie verzichtet werden muss.
Es sei hier nur an die Tatsache erinnert, dass der Mensch nicht deswegen das ultraviolette Licht nicht sehen kann, weil er von seiner chemischen Ausstattung dazu nicht fähig wäre, sondern weil dieses kurzwellige Licht in den transparenten Strukturen seiner Augen Streueffekte erfahren würde, die zu einem verschwommenen, unscharfen und daher auch nicht verwertbaren Bild führen müssten.
Die Komplexaugen der Insekten unterbinden diesen Effekt bekanntlich und die Erfahrungen der Tierphysiologie zeigen denn auch, dass sie das ultraviolette Licht nutzen können.
Umweltbeziehungen
Schon bei der Erörterung der Funktionen von Autos konnte ja gezeigt werden, dass erst einmal eine Unmenge innerer Bedingungen erfüllt sein muss, ehe überhaupt an ein Agieren in einer bestimmten Umwelt gedacht werden kann. Dies gilt ebenso und in noch stärkerem Maße für Lebewesen. Auch sie müssen zuerst einmal friktionsfrei funktionieren, also so strukturiert sein, dass sie sich nicht selber zerstören und die zugeführte Energie auf bestmögliche Weise verwerten und, bei Überschuss, auch entsorgen.
Umweltbeziehungen können grundsätzlich nur vom Organismus aus aufgenommen werden. Ein bestimmtes Lebewesen kann entweder in einer bestimmten Umwelt, in einer ökologischen Nische existieren oder nicht. Wenn nicht, ist es für stammesgeschichtliche Anpassung, die nach Darwin und Wallace Generationen brauchen würde, zu spät.
Lebensräume werden also von den Organismen immer aktiv erschlossen. Die Möglichkeiten der Erschließung hängen von ihren jeweiligen Möglichkeiten ab, die durch die Konstruktion gegeben sind. Niemals konnte Umwelt einen im mechanistisch-darwinistischen Sinne formenden Einfluss auf die Organismen ausüben. Es gibt daher keine stammesgeschichtliche Umweltanpassung, sondern nur eine Eigenentwicklung des Lebens, bei der Umwelt ständig aktiv und immer wieder neu erschlossen wird (Edlinger, Gutmann & Weingarten 1991).
Damit erledigt sich einerseits nicht nur die Anpassungsvorstellung des Darwinismus, ohne dass auch die Evolution der Organismen als Begründung für Lebensfähigkeit und Vielfalt aufgegeben werden müsste. Andererseits kann das Problem der Erkenntnisfähigkeit nun auf der Grundlage der Organismischen Konstruktionslehre neu aufgeworfen und vielleicht auch einer Lösung nähergebracht werden.
Die Evolution
Auf dieser Basis kann die Evolution, der stammesgeschichtliche Wandel der Lebwesen, neu rekonstruiert werden. Er besteht prinzipiell im Wandel energiewandelnder, spontan agierender und weitgehend autonomer hydraulischer Konstruktionen. Die können sich Schritt für Schritt, prinzipiell nur über voll lebens- und funktionsfähige Zwischenstadien abwandeln, wobei die möglichen Abwandlungswege und –bahnen durch die Stammkonstruktionen limitiert sind.
In den letzten Jahren wurden zahlreiche Modelle erarbeitet, die schließlich zu einem generellen Schema des organismischen Wandels mit Schwerpunkt auf der Evolution der Tiere zusammengefasst werden konnten.
Im Unterschied zu den herkömmlichen stammesgeschichtlichen Schemata basiert es auf einem naturwissenschaftlich stringenten Organismusmodell, das es erlaubt, die relevanten Teilbereiche der Biologie wie Morphologie, Physiologie, Genetik u. a. problemfrei zu integrieren. Dadurch zeichnet es sich durch naturwissenschaftliche Stringenz und durchgehende Diskutierbarkeit aus. Jede Kritik, die nicht nur legitim, sondern zum weiteren Erkenntnisgewinn notwendig ist, muss aber entweder auf dem Boden der Organismischen Konstruktionslehre erfolgen oder eine andere theoretische Basis präsentieren, welche diese an Stringenz übertrifft.
Zu sehr ähnlichen biomechanischen Modellvorstellungen und Theorien der Formbildung vom Organismus kam übrigens auch die Schule um Donald Ingber in New York (Ingber 1998).
Rein morphologisch ausgerichteter Formenvergleich oder ausschließlich molekularbiologisch begründete Rekonstruktionen mögen einen gewissen heuristischen Wert haben, den Status von unhintergehbaren Begründungen können sie aber nicht erheben.
Interessant in diesem Zusammenhang ist allerdings, dass auf molekularbiologischen Vergleichsmethoden erarbeitete Verwandtschaftsschemata der letzten Jahre in vieler Hinsicht die durch das Evolutionsschema der Organismischen Konstruktionslehre geforderten stammesgeschichtlichen Abläufe und Verwandtschaftsverhältnisse zwischen den Großgruppen des Tierreichs wiedergeben (s. z. B. Campbell & Reece 2003).
 Abb. 6) Die Evolution des Tierreichs auf der Basis der Organismischen Konstruktionslehre. Diese Evolution ist in die Biosphäre und einen kosmischen Rahmen integriert.
Abb. 6) Die Evolution des Tierreichs auf der Basis der Organismischen Konstruktionslehre. Diese Evolution ist in die Biosphäre und einen kosmischen Rahmen integriert.
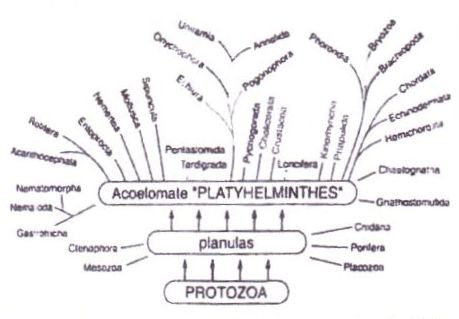
Abb. 7) Im Gegensatz zu den verständlichen und auch kritisch hinterfragbaren Ableitungen der Organismischen Konstruktionslehre präsentieren sich Ableitungen alten Stils als letztlich schlecht überprüfbare, auf willkürlich ausgewählten Merkmalen basierende Schemata, die offen lassen, wie die Vorläufer der rezenten Arten bzw. ausgestorbener Lebensformen beschaffen gewesen sein dürften (Nach Grasshoff, umgezeichnet).
Ausblick
Die vorliegende Publikation ist nur eine in einer ganzen Reihe von Arbeiten die sich kritisch mit dem Darwinismus auseinandersetzen und versuchen, mögliche Alternativen zu ihm aufzuzeigen, ohne an wissenschaftlicher Stringenz und Seriosität zu verlieren. Sie erhebt nicht den Anspruch, fertige Lösungen für alle Probleme im Zusammenhang mit der Evolutionsbiologie zu präsentieren, versteht sich aber als Anregung zu weiterer Diskussion und zu ständiger Überprüfung scheinbar evidenter Sichtweisen. In diese Überprüfung sollen selbstverständlich die hier vertretenen Thesen einbezogen werden. Dies wird umso mehr Ertrag bringen, je weniger die Auseinandersetzung von dogmatischen Immunisierungsversuchen und je mehr sie von echtem Erkenntnisinteresse getragen ist.
Von Konrad Lorenz stammt die ermutigende Aufforderung, sich jeden Tag von einer liebgewordenen Denkgewohnheit zu trennen. Dem ist hier nichts mehr hinzuzufügen.
Literatur
Benderoth, G., Edlinger, K. & Gutmann, W. F. (1995): Lebende Organisation im Rahmen der Megamaschine. – In: Gutmann, W. F. & Weingarten, M. [Hrsg.]: Die Konstruktion der Organismen. II. Struktur und Funktion. – Aufsätze u. Reden Senckenb. Naturf. Ges.,43: 149-178; Frankfurt/M.
Bowler, P. (2003): Evolution: The History of an Idea. – Berkeley / Los Angeles / London: Univ. of California Press.
Campbell, N. A. & J. B. Reece (2003): Biologie. – Heidelberg/Berlin: Spektrum Akademischer Verlag.
Darwin, C. R. (1859): On the Origin of Species by Means of Natural Selection, 2 Vol. – London: Murray. (dt. 1963, Die Entstehung der Arten durch natürliche Zuchtwahl. – Stuttgart: Ph. Reclam u. 1992, Über die Entstehung der Arten durch natürliche Zuchtwahl. – Darmstadt: Wiss. Buchges.).
Darwin, Ch. R. (1868): The Variation of Animals and Plants under Domestication. 2 Vol. – London: Murray. (dt. 1873 u. 1887, Stuttgart: Schweizerbartsche Verlags-handlung.
Edlinger, K. (Ed.) (1989a): Form und Funktion – Ihre stammesgeschichtlichen Grundlagen. Wiener Universitäts-Verlag.
Edlinger, K. (1989b): Sekundäre Wurmkonstruktionen bei amphineuren Mollusken.- In: Edlinger, K. (Hgb.): Form und Funktion – Ihre stammesgeschichtlichen Grundlagen. Wiener Universitätsverlag.
Edlinger, K. (1989c): Zur Evolution der Schneckenkonstruktion l.- Torsion und Frühevolution der Prosobranchier.-Natur und Museum 119(9), 272-293, Frankfurt.
Edlinger, K. (1991a): The mechanical constraints in mollusc constructions – the function of the shell, the musculature, and the connective tissue.- In: Schmidt-Kittler, N. and K. Vogel (Eds.): Constructional morphology and evolution. Springer Berlin-Heidelberg-New York. p. 359-374.
Edlinger, K.(1991b): Die Evolution der Scaphopodenkonstruktion.- Natur u. Museum. 121(4), 116-122.
Edlinger, K. (1992a): Nervensysteme als integrale Bestandteile der mechanischen Konstruktion. – Aufs. u. Reden Senckenb. Naturf. Ges. 39, 131-155.
Edlinger, K. (1992b): Die „Unsichtbare Morphologie“ – Energiefluß und Konstruktion. – Aufs. u. Reden Senckenb. Naturf. Ges. 39, 27-49.
Edlinger, K. (1994a): Das Spiel der Moleküle – Reicht das Organismusverständnis des molekularbiologischen Reduktionismus? – Natur u. Museum 124(6), 199-206.
Edlinger, K. (1994b): Morphologische Determinanten bei der Bildung von Nervensystemen. – Biol. Zent.bl. 113, 25-32.
Edlinger; K. (1994c): Ontogenetische Mechanismen in Beziehung zur Evolution.- In: Gutmann, W. F., D. Mollenhauer und D. St. Peters (Hgb.): Morphologie & Evolution. W. Kramer, Frankfurt/M., 365-384.
Edlinger, K. (1995a): Organismen: Genarrangements oder Konstruktionen? – Eine Replik auf E. Mayr und die Synthetische Theorie. Biol. Zentralblatt 114, 160-169.
Edlinger; K. (1995b): Die Evolution der Plathelmintenkonstruktion – Zur Stammesgeschichte und Systematik der Plattwürmer. – Natur u. Museum 125(110), 305-320.
Edlinger, K. (1998): Konstruktionsprobleme der Miniaturisierung im Tierreich Zur Unumkehrbarkeit konstruktiven Wandels. Natur u. Museum 128(5), 139-151.
Edlinger, K. (2000a): Evolution und Integration lebender Systeme: Aggregation oder Binnendifferenzierung? – In: Edlinger K., W. Feigl & G. Fleck (Hgb.): Systemische Perspektiven. – P. Lang – Europ. Verlag der Wissenschaften., 51-73.
Edlinger, K. (2000b): The Evolution of the mollusc construction: Living organims as energy-transforming systems. – In: D. S. Peters & M. Weingarten (Eds.): Aspects of evolutionary theories. – Senckenberg-Publ. Frankfurt/M. 199-217.
Edlinger K. (2004, im Druck): Charles Darwin und der Maistream der Biologie. – In: W. Feigl, G. Fleck & K. Edlinger (Hrgb.): Jenseits des Mainstreams. Alternative Ansätze von Biologie und Medizin. – Frankfurt/M. / Berlin / Bern / Bruxelles / New York / Oxford / Wien: P. Lang – Europ. Verlag der Wissenschaften, 16-60.
Edlinger K. (2004, im Druck): Alle Evolution ist Anpassung – Ist alle Evolution Anpassung? Eine Kritik der Darwinschen Anpassungsmetapher. – In: G. Fleck, K. Edlinger & W. Feigl (Hrgb.): Die Theorie der Anpassung in Psychologie und Biologie. Frankfurt/M. / Berlin / Bern / Bruxelles / New York / Oxford / Wien: P. Lang – Europ. Verlag der Wissenschaften.
Edlinger, K. and W. F. Gutmann (1997): Molluscs as evolving constructions: necessary aspects for a discussion of their phylogeny. – Iberus 15(2), 51-66.
Edlinger, K. & W. F. Gutmann (2002): Organismus – Evolution – Erkenntnis. Grundzüge der Kritischen Evolutionstheorie und der Organismischen Konstruktionslehre. – Frankfurt/M. / Berlin / Bern / Bruxelles / New York / Oxford / Wien: P. Lang – Europ. Verlag der Wissenschaften.
Edlinger, K., W. F. Gutmann & M. Weingarten (1991): Evolution ohne Anpassung, Verl. W. Kramer, Frankfurt/M.
Gould, St. J. & R. Lewontin (1979): The spandrels of San Marco and the panglossian Paradigm. – Proc. Roy. Soc. London B, 205 No. 1161, 581-598 (http://www.aaas.org/spp/dser/evolution/history/spandrel.shtml).
Grasshoff, M. (1981): Arthropodisierung als biomechanischer Prozess und die Ent-stehung der Trilobiten-Konstruktion. – Paläontologische Zeitschrift, 55 (3/4): 219-235.
Grasshoff, M. (1992a): Die Evolution der Schwämme. I. Die Entwicklung des Kanalfil-tersystems. – Natur und Museum, 122: 201-210.
Grasshoff, M. (1992b): Die Evolution der Schwämme. II. Bautypen und Verein-fachungen. – Natur und Museum, 122: 237-247.
Grasshoff, M. and Gudo, M. (1998a): Die Evolution der Coelenteraten I. Gallertoid-Korallen und Oktokorallen. – Natur und Museum, 128 (5): 129-138.
Grasshoff, M. and Gudo, M. (1998b): Die Evolution der Coelenteraten II. Solitäre und koloniale Polypen. – Natur und Museum, 128 (10): 329-341.
Grasshoff, M. and Gudo, M. (2002): The Origin of Metazoa and the Main Evolutionary Lineages of the Animal Kingdom – The Gallertoid Hypothesis in the Light of Modern Research -. – Senckenbergiana lethaea, 82 (1): 295-314.
Gudo, M. (2002): The development of the critical theory of evolution: The scientific career of Wolfgang F. Gutmann. – Theory of Biosciences, 121 (1): 101-137.
Gudo, M. (2004): Ziele der Evolutionsforschung: Rekon-truktion organismischer Wandlung als Morphoprozess. – In: Feigl, W. G. Fleck & K. Edlinger: Jeseits des Mainstreams. Alternative Ansätze in Biologie und Medizin. – Frankfurt/M. / Berlin / Bern / Bruxelles / New York / Oxford / Wien: P. Lang – Europ. Verlag der Wissenschaften., 61-90.
Gudo, M. and Grasshoff, M. (2002): The Origin and Early Evolution of Chordates: The ‚Hydroskelett-Theorie’ and New Insights Towards a Metameric Ancestor. – Sencken-bergiana lethaea, 82 (1): 325-346.
Gutmann, W. F. & K. Bonik (1981): Kritische Evolutionstheorie. Ein Beitrag zur Überwindung altdarwinistischer Dogmen. – Hildesheim: Gerstenberg.
Gutmann, W. F. & K.Edlinger (1991a): Die Biosphäre als Megamaschine – Ökologische und paläo-ökologische Perspektiven des Konstruktionsverständnisses der Organismen l.- Natur u. Museum 121(10), 302-311.
Gutmann, W. F. & Karl Edlinger (1991b): Die Biosphäre als Megamaschine – Ökologische und paläo-ökologische Perspektiven des Konstruktionsverständnisses der Organismen ll.- Natur u. Museum 121(12), 401-410.
Gutmann, W. F. & K. Edlinger (1994a): Morphodynamik und Maschinentheorie: Die Grundlage einer kausalen Morphologie. – In: Gutmann, W. F., D. Mollenhauer und D. St. Peters: Morphologie & Evolution. W. Kramer, Frankfurt/M.,177-199.
Gutmann, W. F. & K. Edlinger (1994b): Neues Evolutionsdenken: Die Abkoppelung der Lebensentwicklung von der Erdgeschichte.- Archaeoteryx 12, 1-24.
Gutmann, W. F. & K. Edlinger (1994c): Organismus und Evolution Naturphilosophische Grundlagen des Prozeßverständnisses. – In: Bien, G. u. J. Wilke (Hrsg.): Natur im Umbruch – Zur Diskussion des Naturbegriffs in Philosophie, Naturwissenschaft und Kunsttheorie. – Frommann-Holzboog, 109-140.
Gutmann, W. F. & K. Edlinger (1994d): Molekulare Mechanismen in kohärenten Konstruktionen. – In: W. Maier u. Th. Zoglauer (Hrsg.):Technomorphe Organismuskonzepte – Modellübertragungen zwischen Biologie und Technik. Frommann-Holzboog 128, 174-198.
Gutmann, W. F. & K. Edlinger (2002): Organismus und Umwelt – Zur Entstehung des Lebens, zur Evolution und Erschließung der Lebensräume. – Frankfurt/M. / Berlin / Bern / Bruxelles / New York / Oxford / Wien: P. Lang – Europ. Verlag der Wissenschaften
Heikertinger, F. /1954): Das Rätsel der Mimikry und seine Lösung. Eine kritische Darstellung des Werdens, des Wesens und der Widerlegung der Tiertrachthypothesen. – Jena: VEB Gustav Fischer Verl.
Ingber, D. (1998): Architekturen des Lebens. – Spektrum d. Wiss. 3, 32-40.
Jahn, I. (Hrgb.) (2000): Geschichte der Biologie. Theorien, Methoden, Institutionen, Kurzbiographien. – Heidelberg / Berlin: Spektrum Akadem. Verl.
Peters, D. S. & W. Peters (1997): Anpassung – Kernpunkt oder Mißverständnis der Evolutionstheorie? – In: König, V. & H. Hohmann (Hgb.): Bausteine der Evolution. Symposium Übersee-Museum Bremen / Gelsenkirchen / Schwelm: Edition Archaea, 73-82.
Reichholf, J. (1993): Der Schöpferische Impuls. – München: DVA.
Reichholf, J. (2004): Ist die Darwinsche Anpassung nur das Oberflächengekräusel der Evolution? In: Feigl, W. G. Fleck & K. Edlinger: Jeseits des Mainstreams. Alternative Ansätze in Biologie und Medizin. – Frankfurt/M. / Berlin / Bern / Bruxelles / New York / Oxford / Wien: P. Lang – Europ. Verlag der Wissenschaften., 118-141.
Riedl. R. (2003): Riedls Kulturgeschichte der Evolution. – Die Helden, ihre Irrungen und Einsichten. – Berlin / Heidelberg / New York / Hongkong / London / Mailand / Paris / Tokio: Springer.
Wallace, A. R. (1855): On the Law that has Regulated the Introduction of New Species. – Annals and Magazine of Natural History 16 (2), 184-196. (dt. 1870 von A. B. Meyer, Erlangen: E, Besold.
Wallace, A. R. (1858): On the Tendency of Varieties to Depart Indefinitely From the Original Type – Journal of the Proceedings of the Linnean Society: Zoology 3(9): 53-62. (dt. 1870 von A. B. Meyer, Erlangen: E. Besold.